

У популярній культурі кажани огорнуті ореолом таємничості й небезпеки. А насправді це безневинні й вразливі створіння, що потребують нашого захисту. Але щоб захищати, потрібно розуміти, а щоб розуміти – вивчати. Кажанів віддавна досліджують в Інституті зоології ім. І.І. Шмальгаузена НАН України. Частина таких досліджень є глибоко фундаментальними, теоретичними. Та без них ми не матимемо уявлення про те, як рукокрилі живуть і як та чому еволюційно змінюються. Торік добіг кінця проєкт, що його виконували в Інституті за ґрантом Національного фонду досліджень України. У фокусі уваги науковців була еволюція ссавців, що перейшли від наземного способу життя до водного (як китоподібні) та повітряного (як кажани). Про китову частину проєкту ми вже розповідали – настав час поговорити про кажанячу. Слово – завідувачеві відділу еволюційної морфології Інституту зоології ім. І.І. Шмальгаузена НАН України докторові біологічних наук Ігорю Дзеверіну.

Доктор біологічних наук Ігор Дзеверін. Фото Назарія Мазилюка. Джерело: www.pravda.com.ua
– Пане Ігоре, щоквітня у світі відзначають Міжнародний день вшанування кажанів. Звісно, всіх живих істот потрібно шанувати, та й неживу природу – теж. Але чим цікаві саме кажани? Чому їх варто шанувати особливо?
– У кажанів є безліч цікавих рис, які заслуговують на увагу. Починаючи з того, що вони вміють літати, здатні до ехолокації, мають несподівано велику тривалість життя. Взагалі-то у ссавців (та й в інших тварин, але у ссавців це доволі помітно) тривалість життя чітко узгоджена з розмірами тіла. Тобто, наприклад, слони живуть явно довше, ніж миші, а миші – ніж землерийки.
– Існує прямо пропорційна залежність?
– Не те щоб пропорційна, але доволі чітка. А кажани з цього ряду випадають – вони живуть значно довше, ніж можна було б очікувати, з огляду на їхні розміри. Це несподівана риса, і у кажанів їх таких безліч. Водночас, кажани – це абсолютно типові ссавці. За багатьма ознаками вони нічим принципово не відрізняються від мишей, чи зайців, чи слонів, чи людей. Отже, з одного боку, вони типові, з іншого – унікальні.

Кажани виду велика нічниця. Малюнок Ауґуста Шпехта. Джерело: commons.wikimedia.org
Я все життя працюю переважно з кажанами роду нічниць, вони завжди були головним об’єктом моїх досліджень. І дуже цікавим. Передусім рід нічниць надзвичайно різноманітний – він налічує понад 100 видів (для ссавців це дуже багато). Нічниці поширені майже на всьому суходолі. Не можуть жити вони тільки в полярних регіонах типу Арктики й Антарктиди. І ще не дійшли до Нової Зеландії та деяких інших океанічних островів.
– Щодо полярних регіонів зрозуміло – бо там немає комах. А чому нічниці не дісталися до островів? Задалеко?
– Так, через велику відстань від Австралії. Тому в Новій Зеландії живуть усього два види кажанів: один вид гладконосів (це порівняно близькі родичі нічниць) і ще один абсолютно унікальний вид – новозеландський футлярокрилець (Mystacina tuberculata). Чим цікавий цей другий вид? По-перше, містацина (називатимемо її так для зручності) дуже естетично приваблива. Мені здається, її цілком можна було б використовувати для реклами унікальної природи Нової Зеландії – нарівні з ківі й іншими унікальними тваринами. Це лісова тварина, яка ходить по землі, ловить комашок і черв’ячків, добре вміє рити землю. Вона не літає. Точніше, літати вона вміє і чудово зможе злетіти, якщо її сполохати. Але зазвичай містацина польотом не користується. Тільки у разі серйозної небезпеки. Для кажанів це абсолютно нетипово, бо вони здебільшого погано пересуваються по землі і покладаються на політ. Цікаво, що для багатьох птахів цілком нормально більше покладатися на наземну локомоцію (згадаймо курей). Є також чимало видів, вторинно не здатних до польоту, але серед кажанів такого немає. Тобто оця новозеландська містацина – чи не єдиний у сучасній фауні кажан, хто віддає перевагу наземній локомоції. Певний час науковці вважали, що це пов’язано з тим, що у Новій Зеландії майже немає наземних хижаків, тож і багато птахів там теж переходять до наземної локомоції і втрачають здатність літати. Та, вочевидь, із містациною все складніше, бо палеонтологи знайшли її викопних предків з Австралії і з’ясували, що ті теж уміли літати, проте переважно пересувалися по землі. Тобто не у Новій Зеландії річ.

Новозеландський наземний кажан. Фото: Nga Manu Images NZ/Flickr

Містацина народжена не лише повзати. Фото Девіда Маджа. Джерело: www.edgeofexistence.org
Зрештою, так чи так, але містацини дісталися до Нової Зеландії, а нічниці – ні. Проте нічниці є в Європі, Азії, Африці, Америці, Австралії. Як я вже сказав, вони дуже різноманітні. І не зовсім зрозуміло, як такому абсолютно типовому представникові кажанів це вдалося. Нічниці також дуже схожі між собою. А проте можуть використовувати три адаптивні стратегії. Частина нічниць полюють на комах на субстратах – на поверхні землі, на корі дерев, на листі. Як виглядає цей стиль полювання? Кажан летить, бачить комаху на землі, пікірує і ловить її видовженими щелепами. Другий варіант – ловитва комах у польоті. До нього вдаються дрібні нічниці, які дуже красиво літають. І третій варіант – це полювання на комах і навіть на мальків риб біля поверхні води.

Велика нічниця і коник. Фото Мерліна Татла. Джерело: scitechdaily.com

Нічниця ловить нічницю. Кажан одного з північноамериканських видів нічниць – бура нічниця (Myotis lucifugus) – полює на метелика-совку. Фото Майкла Дарема. Джерело: www.batcon.org

Риболовля ставкової нічниці. Фото Глена Томпсона. Джерело: glennsphotos.com
– Я гадала, кажани живляться тільки комахами.
– У принципі, Ви не дуже помиляєтеся. Бо переважна більшість кажанів і, зокрема, переважна більшість нічниць саме комахами й живляться. Але найбільші кажани вже стають рослиноїдними або переходять до живлення дрібними хребетними. Є ціла група справжніх хижаків. Наприклад, серед нічниць є такий вид – пізонікс (українською мовою – «рибоїдна нічниця»), що мешкає в Центральній Америці. То він майже повністю перейшов до живлення рибою. Проте його стратегії полювання цілком відповідають еволюційним стратегіям нічниць, які полюють біля поверхні води.
Отже, повторю, нічниці використовують три різні стратегії полювання. Раніше носіїв цих стратегій вважали підродами в межах роду нічниць. А зараз виявилося, що це екоморфи [життєві форми], які прямо не пов’язані з еволюцією. Тобто кожна з цих трьох екоморф виникала в еволюції нічниць кількаразово, хоча певна спорідненість може спостерігатися. Крім того, в процесі еволюції стратегія могла змінитися. До речі, еволюція екоморф – це популярна тема у дослідників нічниць. Ми з колегами також про це писали. Еволюції екоморф нічниць присвячені, наприклад, моя одноосібна стаття 1998 року, наша спільна з Марією Гхазалі стаття 2010 року [Марія Гхазалі – старший науковий співробітник відділу еволюційної морфології Інституту зоології ім. І.І. Шмальгаузена НАН України, кандидат біологічних наук], а також стаття 2017 року, яку ми з Марією написали й опублікували у співавторстві з бразильським науковцем Рікарду Морателлі [Рікарду Морателлі – таксономіст і біолог дикої природи, дослідник із Фундації Освальду Круша (Ріо-де-Жанейро), доктор наук].
Взагалі отакі роди, що налічують дуже багато видів, які дуже схожі між собою, ідеально годяться саме для еволюційних досліджень. Бо порівнюючи настільки схожі види, можна приблизно зрозуміти, як відбувається еволюція. Про це свого часу писав іще Чарлз Дарвін у праці «Походження видів».
– А в чому тут секрет?
– А ось у чому: види великих і поліморфних родів є мінливішими, ніж види малих родів тих самих родин та більш схожими один на одного, у чомусь нагадуючи, таким чином, різновиди. Ці особливості можна пояснити неповним завершенням процесу видоутворення, тобто ми бачимо початковий етап дивергенції. Насправді у таких родах, як нічниці, ми можемо побачити різні етапи дивергенції: від дуже близьких видів до видів, настільки віддалених, наскільки це взагалі можливо в межах одного роду.
– А якщо види близькі, то можна простежити проміжні етапи?
– Так, у цьому випадку ми бачимо всі проміжні стадії. Маємо можливість побачити, як поступово відбувається дивергенція. Саме тому групи на кшталт нічниць дуже зручні для аналізу еволюції. Минулого разу ми з Вами говорили про масштабне дослідження, в межах якого вивчалась еволюція пристосування до морського середовища на прикладі всього ряду китоподібних. Це дуже масштабна еволюційна подія, але ж вона складається з великої кількості маленьких кроків. І якщо ми хочемо збагнути механізми еволюції, то іноді маємо озброюватись не тільки біноклем чи підзорною трубою, а й поглянути в лупу, а то й у мікроскоп на якісь елементарні, дрібні еволюційні події, на відмінності між близькими видами, а може, і всередині видів.
Мені, як і більшості зоологів, подобається мій основний об’єкт досліджень. Проте він має і об’єктивну цінність – для розуміння еволюційних процесів. Вивчати нічниць у цьому плані теж дуже цікаво. Я маю багато публікацій про нічниць, про різні аспекти їхньої еволюції. А коли зайнявся проблемами інтеграції та модульності, то вирішив подивитись, як вони еволюціонували саме у цій групі. Основним об’єктом стала група нічниць, до якої належать такі поширені у нас види, як велика нічниця і гостровуха нічниця. Це доволі чималі нічниці: за розміром велика нічниця посідає друге місце серед європейських кажанів і перше – серед нічниць, а гостровуха нічниця – третє місце серед європейських кажанів і друге – серед нічниць. Велика і гостровуха нічниці – спеціалізовані представники першої з екоморф, тобто вони полюють на комах (причому великих) із субстратів, а саме – з поверхні землі. Гостровуха нічниця віддає перевагу прямокрилим, її типова здобич – коники та цвіркуни. Велика нічниця теж може ловити цвіркунів, а ще – більших комах із твердішими покривами, наприклад турунів.

Велика нічниця. Фото Жиля Сан Мартена. Джерело: www.flickr.com/photos/sanmartin

Гостровуха нічниця. Автор фото: Адріа Лопес-Бауцельс. Джерело: www.adriabaucells.com
Не менш важливо й те, що є великі наукові колекції зразків нічниць саме цих видів. Фахівці розрізняють варіаційну й еволюційну модульність: обидві форми модульності потребують істотно різних методів дослідження. Щоб дослідити еволюційну модульність (тобто те, чи узгоджено ознаки змінювались еволюційно), вистачить кількох особин одного виду, а от уже для дослідження інтеграції та варіаційної модульності на рівні виду потрібні дуже великі вибірки. Між тим, дослідження варіаційної модульності важливі не менше, якщо не більше, ніж дослідження еволюційної модульності. І це одна з основних причин, що можуть гальмувати подібні дослідження, бо складно зібрати колекції одразу багатьох видів. А у мене є такі вибірки для гостровухої нічниці (Myotis blythii) і великої нічниці (Myotis myotis). Матеріал у таких дослідженнях розподіляють за так званими «операційними таксономічними одиницями» (в моєму дослідженні це щось близьке до підвиду). Сукупно вийшло 5 таких одиниць: велика нічниця з Європи, європейська гостровуха нічниця, близькосхідна гостровуха нічниця, гімалайська гостровуха нічниця (вона так називається, але насправді у мене були збори з Середньої Азії і Казахстану), а також представники унікальної реліктової популяції з Алтаю. Є ще далекосхідний підвид гостровухої нічниці, але великих зборів із нього у мене немає, тому його я не розглядав.
Для порівняння я взяв інший вид нічниць – ставкову нічницю (Myotis dasycneme), еволюційно й екологічно дуже далеку (наскільки це можливо у межах одного підроду) від гостровухої та великої нічниць. Ці ж дві останні настільки близькі, що іноді у них навіть спостерігається міжвидова гібридизація. Це означає, що вони зовсім нещодавно розійшлись еволюційно [тобто розділилися на окремі види]. На відміну від великої та гостровухої, ставкова нічниця має іншу стратегію – як легко здогадатися з її назви, вона є спеціалізованим представником стратегії полювання не з субстратів, як велика та гостровуха нічниці, а з поверхні води.
Моя ідея полягала у тому, щоб на основі порівняння далеких і близьких між собою видів одного роду, а також виокремлення їхніх відмінностей описати інтеграцію та модульність і проаналізувати, як за такого рівня інтеграції ці види відповідають на природний добір. Адже організм – це високоінтегрована система, яка має певні онтогенетичні обмеження на зміни. Види не настільки пластичні, щоб змінюватись одразу, щойно з’явиться адаптивна користь для змін у тому чи тому напрямі. Насправді зміни суттєво залежать від внутрішніх обмежень, тож мені хотілося проаналізувати рівень таких внутрішніх обмежень в обраних видів. Бо хоча про багатьох ссавців і є такі дані, але вони доволі фрагментарні.
Загалом рівень внутрішніх обмежень на зміни тісно пов’язаний із рівнем інтеграції: що вища інтеграція, то вищі ці обмеження (найбільше досліджені в цьому плані особливості черепа, тому те, про що я розповідатиму, стосуватиметься саме черепа). Але є і певні відхилення. А крім того: що вища інтеграція, то нижча модульність. Дуже високою є інтеграція (знов-таки, інтеграція черепа), наприклад, у хижих звірів із родини собачих – собак і вовків. І це зрозуміло: щелепа, зубний апарат – їхній основний «робочий інструмент», а високий рівень інтеграції цього функціонального модуля потрібен для ефективного полювання. Ще вищий рівень інтеграції – у сумчастих, і для них це принципово важливо. Чому? Через особливості розвитку: одразу після народження дитинча має знайти материнські соски, присмоктатися і триматися за них. Якщо зірветься – загине. Тому вже в новонародженого має бути високорозвинений череп із дуже високим рівнем інтеграції. Як, наприклад, в опосума. Причому, можливо, саме це заважає йому еволюціонувати. Бо, як я вже говорив, висока інтеграція черепа обмежує можливості для змін. З іншого боку, опосуми надзвичайно добре пристосовані до середовища, попри те, що не дуже змінилися за останні 70–80 мільйонів років. Ці тварини – класичні «живі копалини».

Опосум. Джерело: owl-online.org
У копитних можливі різні варіанти. Так само – у приматів: наприклад, череп павіана інтегрований майже як у хижих звірів, а у людини й у людських родичів, навпаки, дуже низький рівень інтеграції, висока модульність і низькі обмеження на еволюцію черепа. Можливо, це навіть вплинуло на нашу еволюційну долю.
Тут я, напевно, відійду від теми ще далі, ніж уже відійшов. Для науки доволі важливим є, як його називають, принцип пересічності, або ж принцип Коперника. Якщо коротко: все, що з нами пов’язано, не має жодних особливих переваг. Сонце – абсолютно звичайна, пересічна зоря, у Галактиці таких безліч. І наша Галактика – теж абсолютно пересічна галактика. Наша планета схожа на безліч планет у Всесвіті, хай навіть і не кожна планета має умови, сприятливі для зародження життя. Те саме стосується й еволюції: так вийшло, що цивілізацію створили мавпи, але за інших обставин на їхньому місці могли опинитися восьминоги, мурахи чи інші розумні істоти. Я загалом згоден із цим твердженням, та, водночас, гадаю, принцип Коперника в нашому випадку можна трохи обмежити, бо все ж таки є певні особливості, які допомогли створити цивілізацію саме мавпам. Перша й очевидна особливість – це кінцівки, пристосовані до активного впливу на середовище (інакше кажучи, наявність руки). Друга і теж очевидна – збільшення мозку, а це, своєю чергою, передумова для комунікації, соціальності і багато чого іншого. Хоча мозок – це, скажімо так, дуже високовартісна структура, енерговитратна і складна. Його важко розвивати. Погляньмо на ссавців: у них великий мозок завжди формувався як наслідок збільшення розмірів тіла, тобто якщо збільшувалися розміри тіла, то збільшувався і мозок (хоч і повільніше, ніж розміри тіла). До речі, ця алометрична залежність була порушена, коли ссавці щойно з’явилися: найперші ссавці дуже нагадували своїх родичів-теріодонтів [теріодонти – викопні звіроподібні плазуни], але вже тоді було помітно, що мозок у них трішечки більший, ніж у теріодонта тих самих розмірів. Але далі ссавці залишалися на досягнутому рівні розмірів мозку, мозок збільшувався в еволюції у відповідь на збільшення загальних розмірів, причому розмір мозку збільшувався повільніше, ніж розмір тіла. І фактично лише у лінії гомінід (людиноподібних) проявилася протилежна тенденція – тіло почало рости у відповідь на збільшення мозку.
А тепер повернімося трохи назад. У гомінід (якщо розуміти їх у широкому сенсі, включно з шимпанзе і горилою) рівень обмежень на еволюцію розмірів черепа мінімальний серед ссавців. Отже, їхній мозок міг збільшуватися майже безперешкодно, чи, принаймні, з меншими перешкодами, ніж у інших ссавців. Тоді як в інших еволюційних лініях скорельованою відповіддю на збільшення мозку було б дуже суттєве зростання розмірів тіла. Але за такої негативної алометрії [алометрія – нерівномірний ріст частин тіла на різних етапах індивідуального розвитку] значне збільшення розмірів тіла супроводжувалося б незначним збільшенням розмірів мозку. У наших предків такого не було: вся система дуже пластична, і це явно дало їм змогу за лічені мільйони років утричі збільшити мозок.
Порівняймо череп і мозок неандертальця з нашим мозком і черепом: за розмірами вони майже однакові (у неандертальця навіть трішечки більші), а от форма – різна. У нас череп, а отже, й мозок – опуклі, у неандертальця – пласкіші й видовженіші. Вважається навіть, що в ранньому онтогенезі [онтогенез – індивідуальний розвиток організму від запліднення до природної смерті] сучасної людини з’явилася стадія глобуляризації, коли череп набуває своєї опуклої форми. Ця тема сама по собі дуже цікава для антропологів, котрі досліджують, як мало відрізнятися мислення неандертальця за такої форми черепа й мозку, що у нього могло було більш розвинутим, а що – менш розвинутим. Я сформулюю дещо інший аспект цього питання. Сапієнс і неандерталець – дуже близькі види. Настільки, що, можливо, у них спостерігалася міжвидова гібридизація (принаймні в обмежених масштабах). Наш геном містить щонайменше кілька генів, запозичених у неандертальців. До чого я веду? Те, що такі близькі види настільки різняться за формою черепа, є абсолютно нетиповою для ссавців ситуацією. Зазвичай близькі види дуже подібні між собою за цією ознакою. Наприклад, собака, вовк, шакал і койот переважно різняться розмірами, а не формою. А якщо форма почала змінюватися, то вже, вочевидь маємо до діла з різними родами: наприклад, з одного боку – собака, вовк, шакал і койот, а з іншого – лисиця, корсак, фенек і песець; також до окремих родів належать чагарниковий собака, гривастий вовк, гієноподібний собака, єнотоподібний собака. Так само й у кажанів: близькі види різняться, радше, розмірами, ніж формою. Тож якщо форма змінилася на рівні дуже близьких між собою видів, то це означає, що онтогенетичні обмеження на еволюційні зміни зовсім слабенькі.
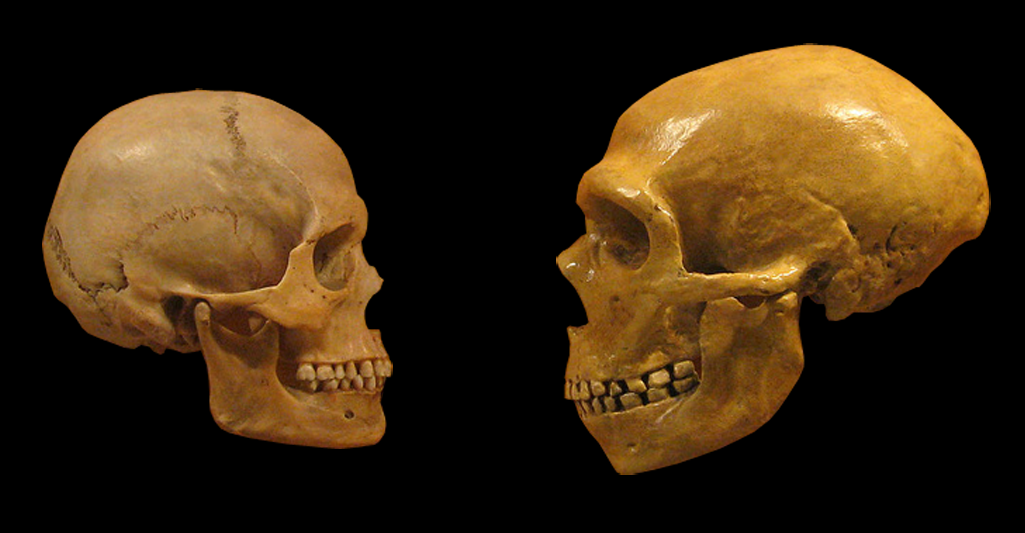
Ліворуч – череп людини розумної, праворуч – череп неандертальця. Джерело: commons.wikimedia.org
– Чи означають ці еволюційні зміни, що й умови середовища змінилися кардинально? Чи у випадку гомінід – не конче?
– Хтозна, як і чому ця зміна сталася. Взагалі-то, неандерталець – це європейський вид, і він пристосований до більш суворих умов (а клімат в Європі тоді був дуже суворий). Сапієнси прийшли з Африки, але теж пристосувалися до цих суворих умов. Зрештою, вижили все ж таки сапієнси, а не неандертальці. А чому – не зовсім зрозуміло. Припускають, що річ у вищому рівні соціальності сапієнсів. Неандертальці жили сім’ями, мало контактували з іншими представниками свого виду, а сапієнси утворювали численніші орди і, можливо, саме завдяки вищому рівню соціальних зв’язків здобули перевагу перед неандертальцями. Попри те, що, з формальної точки зору, неандертальці не лише виглядали пристосованішими до дуже холодного клімату (вони вирізнялися кремезною статурою і були фізично сильнішими), а й мали більший мозок, тобто могли бути розумнішими за наших предків. Хоча насправді ніхто ж не знає, як форма мозку неандертальця впливала на його мислення.
– Ви сказали, що наші предки у найдавніші часи жили ордами. Наскільки це численна зграя, юрмище?
– Вважається, що кілька десятків, але, можливо, й сотні (для порівняння: у неандертальців, вочевидь, було до десяти особин у групі). Але достеменно теж невідомо. Це припущення, що базується на спостереженні за найпримітивнішими сучасними племенами й аналізі палеонтологічних знахідок.
– У природі один вид, буває, витісняє іншого з його ареалу. Чи могло так статись і з неандертальцями?
– Цього ніхто не знає, але така версія виглядає доволі правдоподібною. Що я можу сказати: люди – дуже агресивні істоти. І, судячи з усього, переважна більшість контактів і серед сапієнсів, і серед неандертальців були вбивствами. Конкуренція була дуже жорстка. Як свідчать знахідки, доволі велика частка людей того часу мають ознаки насильницької смерті. Це тема для розмови з палеоантропологами. Гуманізм поставав дуже поступово.
– Вважають, що це здобуток порівняно недавній, останніх кількох тисяч років.
– Я думаю інакше. Мені здається, що він, навпаки, розвивався вже тоді і навіть набагато раніше. Бо без взаємодопомоги маленькій суспільній групі (хай навіть це й орда зі 100 особин) було б дуже важко вижити. Водночас, тип соціальності у сапієнсів (і, не сумніваюся, у неандертальців теж) дуже специфічний – це взаємопідтримка всередині своєї групи й агресія щодо інших груп. Як прогресувала моральність, етика? Межі «своєї групи» розширювались і охоплювали дедалі більшу кількість особин. Спершу це були найближчі родичі, потім зграя, плем’я, далі – вся нація, все людство. Сьогодні ми потроху переносимо свої етичні принципи і на тварин.
Отже, як на мене, то прогрес етики полягав у тому, що люди стали розглядати представників інших племен не як об’єкт для полювання, а як своїх, з якими треба співпрацювати і яких заборонено вбивати. Я думаю, що це було вже у мавп (бо всі мавпи є високосоціальними тваринами), а далі розвивалося, хоч і дуже повільно, як у неандертальців, так і у сапієнсів. І ця риса – разом із деякими іншими – допомогла нашому видові створити цивілізацію.
Теоретично, якщо уявити, що людство раптом кудись зникне, то шанси створити власну цивілізацію з’являться у дуже багатьох видів тварин. Наприклад, у каланів. Чому? По-перше, вони дуже розумні. По-друге, дуже соціальні. По-третє, звикли рухатись у тривимірному просторі, а це дуже сприяє розвиткові мозку. Те саме стосувалось і нас: мавпи теж навчилися рухатись у тривимірному просторі, бо жили на деревах, з яких потім спустилися. По-четверте, калани вміють користуватися примітивними знаряддями – наприклад камінням, за допомогою якого вони розбивають тверді покриви своєї здобичі. Але для того, щоб створити цивілізацію, каланам, як і нашим предкам, доведеться навчитися ходити на задніх ногах і звільнити передні кінцівки для роботи зі знаряддями. Усе це можна уявити, проте у хижих звірів дуже високий рівень онтогенетичних обмежень на еволюцію черепа і мозку. Їм складніше буде збільшити мозок. Хоча навіть такі обмеження може зруйнувати дуже інтенсивний природний добір. Але все одно це додаткові труднощі.

Калан (він же – морська видра). Джерело: commons.wikimedia.org
– Пане Ігоре, повернімося, будь ласка, до кажанів. У межах проєкту за ґрантом Національного фонду досліджень України ви з колегами досліджували модульність, інтеграцію, дезінтеграцію, реінтеграцію організму кажанів під час еволюційних змін при переході з одного середовища в інше. Тобто предки кажанів були наземними тваринами? Хто ці предки і якими вони були?
– Які вони були – не відомо. Найпримітивніші кажани, яких ми знаємо, – це рід Onychonycteris, і це вже на 90% сучасні кажани. Найхарактерніші їхні риси – кігтики на всіх пальцях крил (у сучасних кажанів вони втрачаються) і непропорційно короткі крила, що, втім, не заважало цим тваринам літати (хоч і не дуже добре). Щоправда, вони ще не здатні були до ехолокації. Але будь-хто, хто порівняє їх із кажанами, скаже, що оніхоніктеріси – це кажани.

Оніхоніктеріс у скам’янілостях. Джерело: en.wikipedia.org/wiki/Onychonycteris
– Тобто кажани у палеонтологічному літописі вигулькують ніби нізвідки?
– Фактично, так. Із китоподібними значно простіше.
– А чому так? Минулого разу Ви розповідали, що череп кажанів стоншується, щоб полегшити їхній політ, тому кістяк потім гірше зберігається. Відсутність давніших даних пов’язана з цим?
– Так, гадаю, саме з цим. Бо практично та сама проблема і з птахами. Літопис еволюції птахів такий же фрагментований, як і у кажанів. Хоча про птахів відомо все-таки трішечки більше. Але, сподіваюсь, і предка кажанів колись іще розшукають. Так чи так, завдяки генетичному аналізу вдалося з’ясувати, що кажани є родичами непарнокопитних, хижих звірів і комахоїдних.
– А хто їхні найближчі родичі серед сучасних видів?
– Еволюційні шляхи кажанів та інших ссавців розійшлися доволі рано. Тому найближчих родичів кажанів можна визначити лише на рівні рядів. Це непарнокопитні, хижі звірі та комахоїдні, усі вони – приблизно на одному рівні спорідненості з кажанами. Перші два ряди, можливо, трохи ближчі до кажанів. Для цієї лінії навіть запропоновано назву Pegasoferae, тобто «пегасохижі» (кінь + крило = Пегас; Пегас + хижі = пегасохижі). Але це не така близька спорідненість, як усередині рядів, наприклад, між кіньми, тапірами, носорогами, халікотеріями і так далі серед непарнокопитних або навіть у китоподібних із парнокопитними. До речі, парнокопитні разом із китоподібними є паралельною еволюційною лінією, близькою до лінії пегасохижих. А всі вкупі вони утворюють групу так званих лавразіатеріїв [лавразіатерії – плацентні ссавці, що походять із давнього суперконтиненту Лавразія, яка об’єднувала сучасні Північну Америку, Гренландію, Європу й Азію], пов’язану своєю еволюцією з Північною Америкою. Ця група дала дуже велику кількість різноманітних груп ссавців, хоч і не всіх. Окрему групу становлять афротерії, пов’язані, як випливає з їхньої назви, із Африкою (наприклад хоботні). До ще однієї групи належать, з одного боку – примати, а з іншого боку – гризуни. Є також група неповнозубих [неповнозубі вважаються одними з найпримітивніших серед плацентних ссавців, вирізняються особливостями хребцевих суглобів], центр розселення яких – це Південна Америка. У сумчастих історія взагалі багата на події: вони походять із Північної Америки, але там повністю вимерли, а натомість мігрували до Південної Америки, звідти – до Антарктиди, а з Антарктиди – до Австралії, де дали вибух розселення. У Південній Америці лишилося тільки кілька видів. А потім опосуми посунули назад і знову заселили Північну Америку – як вторинні вселенці. Здається, це чи не єдиний випадок, коли південноамериканський вид заселяє Північну Америку, бо зазвичай відбувалося навпаки, тож видів північноамериканського походження у Південній Америці значно більше (ті ж верблюдові чи ягуари).
Нічниці – не настільки старий рід. Вони відокремилися значно пізніше, коли кажани вже сформувались як група. Свого типового вигляду – з крилами, перетинками й ехолокаційним апаратом – кажани набули дуже швидко і далі майже не змінювалися. Свого часу я взявся перевірити, з чим пов’язаний цей консерватизм – із певними обмеженнями чи тиском середовища (тобто поєднанням живлення комахами з біомеханічними обмеженнями на політ). Це стало однією з основних тем моєї дисертації [мається на увазі дисертація Ігоря Дзеверіна «Механізми трансформації структур черепа в еволюції нічниць та споріднених груп гладконосих рукокрилих» (2012 рік) на здобуття ступеня доктора біологічних наук]. Крім того, у нас із Марією Гхазалі є спільна стаття з цієї теми. Головний висновок полягає у тому, що провідну роль відіграють все-таки вимоги адаптивної зони – вимоги до польоту і до харчування. На низькому таксономічному рівні кажани здатні еволюціонувати доволі швидко, але – у межах своїх екологічних ніш і адаптивної зони. А адаптивна зона не дуже дає змогу, наприклад, збільшувати розміри та форми. До речі, те саме стосується і птахів: вони надзвичайно різноманітні, та все ж масштаб цієї різноманітності значно менший, ніж у ссавців.

Політ нетопира карлика. Фото Рудмера Цверфера. Джерело: www.welcomewildlife.com
– А наскільки споріднені з кажанами так звані летючі собаки й летючі лисиці? Вони ж теж є представниками рукокрилих?
– Колись дуже популярною була ідея дифілії кажанів. Тобто ідея про те, що кажани поділяються на дві окремі групи, два підряди чи навіть незалежні ряди: криланів (тобто летючих собак, летючих лисиць і споріднені форми) і решту (так званих летючих мишей, або ж усіх, хто не є криланами, зокрема це – гладконосі, листоносі й підковоносі). Деякі дослідники навіть вважали, що крилани є родичами приматів, а летючі миші – родичами комахоїдних.
– А як насправді?
– Насправді це припущення не підтвердилося. І морфологічні, і генетичні дослідження показали, що рукокрилі – це єдина цілісна група. Ця група, як свідчать сучасні дослідження, поділяється на дві підгрупи: Yangochiroptera та Yinochiroptera. До першої належать численні групи, зокрема гладконосі та листоносі. До другої – підковоносі та несправжні вампіри. Я припускаю, що розробники цієї класифікації захоплювалися східною філософією, і запропоновані ними назви слід перекладати українською як, відповідно, ян-кажани (або ян-рукокрилі) та інь-кажани (або інь-рукокрилі). Так от, виявилося, що крилани близько споріднені з інь-рукокрилими, вони значно ближчі до підковоносів або несправжніх вампірів, ніж підковоносі – до гладконосих чи листоносих кажанів. Крилани – це лише одна з родин ряду кажанів, а їхні унікальні особливості пояснюються ранньою дивергенцією і пристосуванням до рослинної їжі.

Індійська летюча лисиця. Фото Яна Арендтса. Джерело: www.flickr.com/photos/janarendtsz/
Але повернуся до нічниць. Отже, після того, як ми визначили темпи їхньої еволюції, у мене виникла ідея поглянути, як змінюються модульність і інтеграція організму нічниць, і як ці тварини відповідають на вплив природного добору. Щодо другої частини завдання: загалом відповідь будь-якого організму на певні зміни у середовищі суттєво детермінується його (організму) властивостями. Тобто організм має тенденцію змінюватися в напрямі, який відповідає його внутрішнім онтогенетичним і генетичним обмеженням.
Цей напрям іще називають лінією найменшого еволюційного опору. Висловлюючись математичною мовою, це відповідає першій головній компоненті коваріаційної матриці адитивних генетичних значень ознак. Спробую пояснити так, щоб не заглиблюватись у теорію. Кількісну ознаку можна схарактеризувати її фенотипним значенням. Наприклад, загальна довжина черепа у гостровухої нічниці становить зазвичай від 20 до 23 мм. Фенотипне значення можна розділити на кілька складників, серед яких найважливішим є адитивне генетичне значення. Це та частина фенотипного значення, яка передається спадково. Зазвичай такі ознаки детерміновані великою кількістю генів, для кожного можна порахувати середній ефект, суму цих ефектів становить адитивне значення, тому його і називають адитивним. Для морфометричних ознак адитивне значення зазвичай становить значну частину фенотипного значення; решту становлять наслідки взаємодії між генами та впливу середовища. Оскільки еволюція – це зміна ознак у низці поколінь, для розуміння еволюції насамперед треба працювати саме з адитивними значеннями. Ми можемо оцінювати дисперсію адитивних ознак, їхню кореляцію, можемо визначити той напрям у фазовому просторі ознак, де адитивна дисперсія максимальна. Це і є лінія найменшого еволюційного опору.
Відповідь популяції на добір залежить, з одного боку, від властивостей самого організму, а з іншого боку – від інтенсивності добору. Найкраще добір діє, якщо він збігається з лінією найменшого еволюційного опору. У синтетичній теорії еволюції є узагальнення, яке називають Фішеровою фундаментальною теоремою природного добору. Її суть полягає у тому, що інтенсивність добору пропорційна адитивній генетичній дисперсії за пристосованістю. Іншими словами, найбільше матеріалу для дії добору буде там, де найбільша адитивна генетична мінливість. А це і є лінія найменшого еволюційного опору. Та навіть якщо добір діє в якомусь іншому напрямі, але при цьому він доволі інтенсивний, то все одно зможе змінити популяцію, хай у неї і не така велика мінливість. Отже, відповідь популяції на добір – це певний компроміс між напрямом дії добору і напрямом максимальної мінливості. Що інтенсивніший добір, то ближчою буде відповідь популяції до напряму його дії. Що інтенсивніші онтогенетичні й генетичні обмеження на еволюцію, то ближчою буде відповідь до лінії найбільшої мінливості.
І, що цікаво, випадкова зміна, тобто зміна внаслідок генетичного дрейфу, теж, найімовірніше, відбуватиметься саме в цьому напрямі. А якщо вид змінюється в напрямі найменшого еволюційного опору, то доволі нетривіальним завданням буде визначити – випадкова це зміна внаслідок генетичного дрейфу чи спрямована зміна внаслідок природного добору. Згрубша, можна сказати, що лінія найменшого еволюційного опору – це лінія збільшення або зменшення розмірів. Як я вже говорив, всередині роду види різняться переважно розмірами, а значні відмінності форми вже позначають радикальніші еволюційні зміни – утворення нових родів чи родин.
Моя ідея полягала у тому, щоб визначити інтеграцію в обраних таксономічних одиниць з роду нічниць, з’ясувати рівень їхніх онтогенетичних обмежень і проаналізувати, як ці види еволюціонують. Оскільки гостровуха нічниця вирізняється дуже великими, як для нічниць, розмірами, а велика нічниця – ще більша, то зрозуміло що лінія, до якої належать ці два види, еволюціонувала в бік збільшення розмірів. Це означає, що така група ідеально годиться для дослідження значення лінії найменшого еволюційного опору.
Що ж вдалося дізнатися? Найперше було з’ясовано рівень інтеграції черепа у великої і гостровухої нічниць, а також змодельовано їхню відповідь на добір. Щодо кажанів було не дуже багато даних із цього питання. З’ясувалося, що і рівень обмежень, і рівень інтеграції черепа у великої та гостровухої нічниць залежить від розмірів тіла і від розмірів черепа. Найнижчим він є у ставкової нічниці (нагадаю, що я обрав цей вид для порівняння), вищий – у гостровухої нічниці, ще вищий – у великої нічниці. Тобто череп великої нічниці виявився не лише найбільшим за розміром, а й найінтегрованішим. З огляду на наявні тоді дані, це був доволі нетривіальний результат. Така залежність майже не помітна у раніше досліджених ссавців, але в досліджених мною видів вона виявилася дуже чіткою. Я припустив (це, звісно, гіпотеза), що це пов’язано з особливостями стратегії полювання. Річ у тім, що розмірами різняться не лише нічниці, а й комахи, на яких вони полюють, а також твердість покривів цих комах. Тобто у нічниць розміри черепа чітко корелюють із розмірами і твердістю їхньої здобичі. Що більша нічниця, то більших комах вона здатна ловити. Це перевірено, про це йдеться у нашій спільній із Марією Гхазалі статті 2013 року, на яку я спираюсь у кількох наступних своїх наукових публікаціях – як одноосібних, так і у співавторстві.
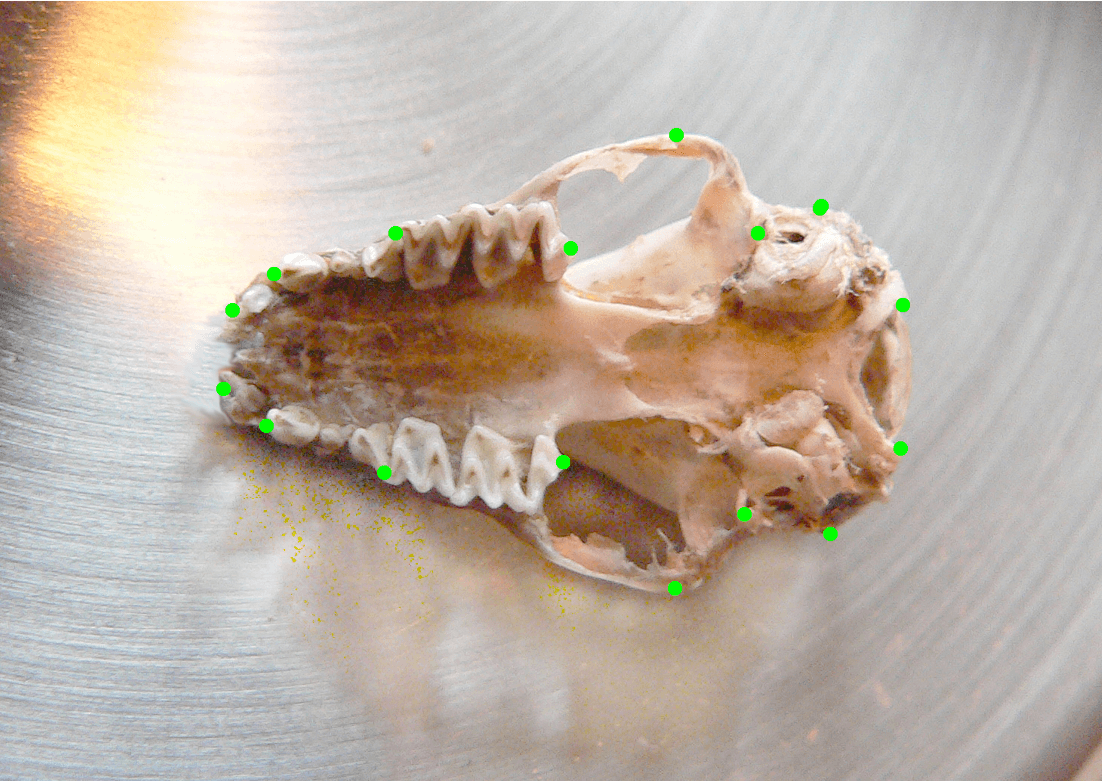
Череп великої нічниці з мітками для геометричної морфометрії. Автор світлини – Ігор Дзеверін. Джерело: https://www.imath.kiev.ua/events/index.php?seminarId=93&archiv=1 (відеозапис: https://youtu.be/AZlS6HRauFs)
Але зовсім невеличкій тварині (хай і доволі крупній порівняно з близькими видами) складно спіймати великого цвіркуна чи коника і втримати його, а спіймати туруна ще складніше, і це потребує високої інтеграції в щелепному апараті. А отже, й череп потрібен інтегрованіший. Нарешті, більша кореляція між ознаками і вищий рівень інтеграції означають також вищий рівень онтогенетичних обмежень.
– Ви сказали, що висунули гіпотезу і шукаєте аргументів на її користь. Отже, Ви схильні вважати, що вона таки підтвердиться?
– Усі дані, які ми маємо на цей момент, дуже добре з нею узгоджуються. За цією ж схемою непогано було б дослідити кінцівки ставкової нічниці, в якої і розміри тіла, і рівень інтеграції черепа виявилися найменшими. Річ у тім, що ставкова нічниця має зовсім іншу стратегію – вона полює на водних комах за допомогою своїх задніх кінцівок, використовуючи їх як своєрідний остень чи щось подібне. Тому логічно очікувати, що її череп буде менш інтегрованим не лише через розміри, натомість вищою буде інтеграція елементів задньої кінцівки. Сподіваюся рано чи пізно виконати таке дослідження. Для нього потрібно більше даних про задню кінцівку ставкової нічниці.
Наразі ж усе сходиться: розміри тіла кажанів і розміри комах, на яких вони полюють, корелюють із рівнем інтеграції черепа кажанів. Це, повторю, абсолютно нетривіальний висновок. Він був головною ідеєю статті 2020 року, яка вийшла друком, коли ми почали працювати над проєктом, поданим потім на конкурс Національного фонду досліджень України. Ми вирішили дослідити цілісність організмів під час переходу видів з одного середовища до іншого, і ці напрацювання лягли в основу нашого проєкту. Було зрозуміло, що за розпочатим напрямом варто працювати далі. А це означало дослідити ще й модульність. Тож, коли ми виграли ґрант від Національного фонду досліджень України, то взялися, зокрема, вивчати модульність у тих само таксономічних одиниць із трьох видів роду нічниць: вже знайомих нам великої нічниці, гостровухої нічниці та ставкової нічниці.
Найперший наш висновок: що вища інтеграція, то менша модульність. Але це найменш цікавий результат, бо він очікуваний, абсолютно тривіальний і стосується майже всіх тварин.
А от що справді цікаво було зрозуміти, то це те, як узагалі ознаки розподіляються за модулями. Бо підходів є багато, зокрема традиційна морфометрія та геометрична морфометрія тут дають дещо різні розподіли [традиційна морфометрія аналізує поздовжні та поперечні розміри, маси, кути, площі, індекси; геометрична морфометрія описує і аналізує топографію структур із використанням дво- чи тривимірних координат]. Якщо добре знати біологію тварин, то, з огляду на їхній спосіб життя, приблизно можна припустити, які ознаки об’єднуватимуться в які функціональні модулі. Тобто спершу можна гіпотетично розподілити ознаки за модулями, а потім – за допомогою спеціального математичного алгоритму перевірити, чи збігаються гіпотетичні модулі з реальними.
– Модулі – це ознаки, які працюють на виконання якоїсь функції?
– Так. Це ознаки, які мають високу кореляцію між собою. Зазвичай це пов’язано з їхнім спільним функціонуванням, але може бути пов’язано також і з їхнім спільним онтогенетичним утворенням. В основі, безперечно, лежить генетична спорідненість, але генетична база адаптована до того, щоб забезпечити функціонування.
– Як багато функціональних модулів у кажанів, яких ви з колегами досліджували? І що ви у ці модулі виокремили?
– У різних ссавців по-різному, адже модульність зазнавала еволюційних змін. До того ж різні методи оцінювання дають дещо різні оцінки модульності. У першому наближенні на черепі кажанів можна виокремити два модулі (мозкова капсула і рострум), але, наприклад, у підковоносів – завдяки розвитку носової лопаті – виокремлюється ще третій модуль. Можливий і більш дрібний поділ. В однопрохідних [яйцекладних] ссавців на черепі два високоінтегровані модулі, а у сумчастих і плацентних можна виокремити 6 модулів різного рівня інтегрованості. Про це цікаво пише Анджалі Ґосвамі – одна з найталановитіших і найвідоміших сучасних дослідниць еволюції. До речі, вона є співавторкою концепції мухи в пробірці, про яку ми говорили минулого разу.
На основі того, що нам відомо про життя нічниць, можна гіпотетично запропонувати для перевірки кілька варіантів поділу ознак їхнього черепа на модулі. Водночас, хотілося б, щоб математичний алгоритм сам шукав і визначав модулі, аналізуючи кореляції між ознаками. Тут я маю зізнатися, що вже багато років живу подвійним життям (усміхається): займаюсь еволюційною біологією в Інституті зоології ім. І.І. Шмальгаузена НАН України й одночасно викладаю чисельні методи у статистиці на кафедрі теорії ймовірностей, статистики та актуарної математики механіко-математичного факультету Київського національного університету імені Тараса Шевченка, цікавлюся статистичними проблемами, статистика – це також моя тема.
– Несподіване поєднання.
– Так. Найпростіший шлях виявлення функціональних модулів у живих організмів за допомогою математичного алгоритму – це кластерний аналіз: будують кореляційну матрицю, визначають, які ознаки корелюють між собою тісніше, і представляють зв’язки у вигляді дендрограми (ієрархічного дерева). Це класична методика, із використанням якої виконано дуже багато робіт. Кластерний аналіз широко застосовується не лише для класифікації ознак за кореляціями, а й для класифікації у біологічній таксономії. Але у звичних алгоритмів кластерного аналізу є суттєвий недолік: ці методики не мають оцінок статистичної значущості в явному вигляді. Тобто класичні моделі кластерного аналізу дають змогу побудувати дендрограму, а з’ясувати, наскільки вона відповідає дійсності, – ні. Проте і цю проблему зараз уже розв’язано. Команда японських дослідників (основний автор – Хідетоші Шимодайра [професор Університету Кіото]) поєднала кластерний аналіз із методикою бутстрепу. Бутстреп – це один із найвпливовіших і найзручніших методів так званого ресемплінгу (українською мовою ресемплінг перекладають як «методи генерації повторних вибірок»), він дуже широко застосовується у сучасній статистиці для перевірки даних. Завдяки чому тепер можна не просто побудувати дендрограму, а й обчислити ймовірність кожного відгалуження цього дерева зв’язків. Японські науковці розробили для цього спеціальний пакет мовою програмування R. З ним дуже зручно і легко працювати, попри те, що сама методика доволі складна. По-моєму, це суттєво поліпшило роботу з кластерним аналізом, ми в Інституті зоології активно користуємося цією методикою.
Але ми спробували й інші варіанти. Наприклад, логіку теорії графів і теорії мереж, бо кластерний аналіз лише допомагає побудувати дерево зв’язків, а не дійти висновку, до якого модуля належить та чи та ознака. Застосовуючи теорію мереж, вузлами ми вважали ознаки, а ребрами – зв’язки між ними. А також намагалися класифікувати ознаки за допомогою дуже поширеного в теорії мереж методу Ньюмена [Марк Ньюмен – британський та американський фізик і математик, професор Мічиганського університету]. Цей метод доволі потужний і дає непогані результати, але, на жаль, теж не оцінює значущість. Тому наша ідея полягала в тому, щоб побудувати такі алгоритми, які оцінювали б імовірність кожної з одержаних груп. Я запросив Оксану Верцімаху – на той час випускницю університету, чия дипломна робота присвячувалася методу Ньюмена (зараз Оксана – працівниця нашого Інституту [Оксана Верцімаха – молодший науковий співробітник відділу еволюційної морфології Інституту зоології ім. І.І. Шмальгаузена НАН України]) взяти на себе розроблення таких алгоритмів, і Оксана блискуче впоралася з цим завданням: розробила математичний алгоритм, який дав нам змогу шукати функціональні модулі у черепі нічниць. Суть алгоритму – в поєднанні пошуку груп за методикою Ньюмена і перевірки результатів (тобто визначення ймовірності об’єднань) за допомогою підходу Шимодайри та співавторів. І цей метод ми випробували на модулях нічниць.
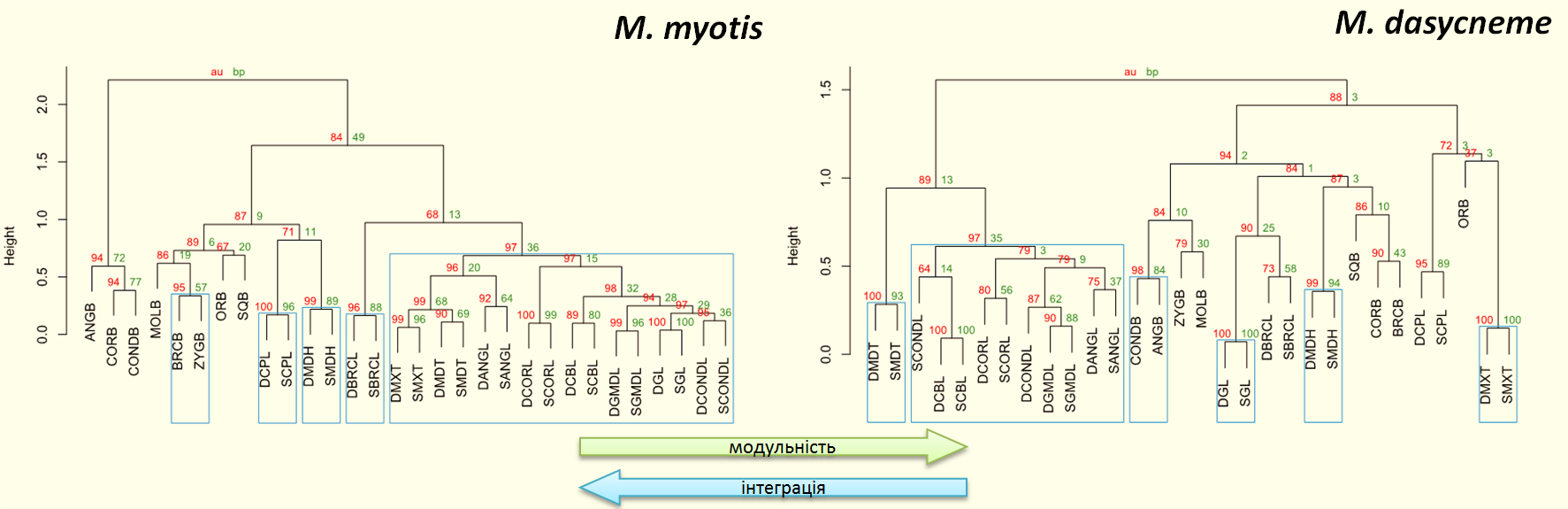
Ієрархічні дерева великої (ліворуч) і ставкової (праворуч) нічниць: кореляції між краніометричними ознаками. У великої нічниці вища інтеграція черепа, у ставкової нічниці вища модульність черепа. Червоний колір – наближено незсунені бутстреп-оцінки (у відсотках), зелений – стандартні бутстреп-оцінки для кожного з кластерів (також у відсотках), рамки – статистично найкраще підтверджені кластери
Що ж вийшло? Результат – непоганий. З-поміж усіх апріорних моделей, які ми перевіряли, найкращою виявилася найпростіша, яка передбачала наявність двох модулів у черепі нічниць: до одного з них потрапили поздовжні проміри, до іншого – поперечні (так само, вибираючи одяг, ми теж зважаємо на зріст і розмір, бо закони морфогенезу в людей і нічниць не дуже різняться). І загалом ця апріорна модель добре узгоджується і з результатами нашого дослідження, і з даними про інших ссавців. Інші запропоновані для перевірки моделі дали гірші результати. Апостеріорна ж модель, побудована математичним алгоритмом, вийшла на рівні найкращої з апріорних, хоч якість все одно не ідеальна. Але ліпшого і не варто було очікувати, тому що класифікація – це все-таки дуже умовна річ. Насправді в організмі зазвичай чимало проміжних ознак, наближених одночасно (хоч і різною мірою) до кількох модулів. Якщо, скажімо, певна ознака належить до одного модуля з імовірністю, що лише трохи більша, ніж імовірність належності до іншого модуля, то ми все одно маємо жорстко віднести її до того чи іншого з цих модулів. Тобто розшукати ідеальну відповідність проблематично. Але можна досягти непоганої, що, здається, в нас і вийшло.
Крім того, наші дослідження показують, що досягти непоганої відповідності можна різними шляхами. Тобто різні класифікації можуть бути однаково точними. Це цікаво для подальшого розвитку машинної класифікації. І, до речі, зовсім не факт, що взагалі потрібно шукати ідеальної відповідності. Необхідні, радше, різноманітні варіанти – залежно навіть не лише від алгоритму, а й від мети, яку ставлять перед собою дослідники. По суті, розмежування ознак за модулями виконує роль мапи. А хороша мапа ж не повторює всіх особливостей ландшафту – вона просто допомагає зорієнтуватися.
Повертаюся до нашого дослідження. Нам довелося вимірювати рівень модульності, а це теж потребувало серйозного математичного апарату. Тож ми з Оксаною знову застосували доволі складну схему. І для обраних таксономічних одиниць показали, що у цій еволюційній лінії – обернена залежність між модульністю і розміром: що менший розмір, то вища модульність. А, відповідно, це ще одне підтвердження того, що у цій лінії збільшення розмірів тіла тягне за собою зростання інтеграції.
Але можна зробити і загальніший висновок. Поясню. Ще раніше я протестував кореляційні системи черепа досліджених видів, щоб зрозуміти, чи є відмінності між ними результатом випадкового генетичного дрейфу чи природного добору. Вийшло, що це дія добору. І цей висновок схожий на те, що побачили наші колеги, досліджуючи китів: кажани еволюціонують під впливом релаксованого природного добору. Гадаю, це взагалі типова ситуація. Випадкові зміни в певному напрямі (генетичний дрейф) можуть бути доволі значними, чи їх може не бути, вони не мають визначеного напряму, тому слабенький, але спрямований природний добір все одно забезпечує загальний напрямок еволюційної зміни. Отже, Фішер і Голдейн [Рональд Фішер і Джон Голдейн – британські науковці, засновники синтетичної теорії еволюції, дотримувалися точки зору, згідно з якою визначальну роль в еволюції відіграє природний добір] таки мають рацію, однак і роль дрейфу в цьому процесі теж дуже значна.
До чого я веду? Мені здається, що і інтеграцію, і модульність потрібно розглядати як такі ж адаптації, як будь-які ознаки самі по собі. У процесі еволюції тварини пристосовуються до умов зовнішнього середовища: якщо клімат холодний – відростає пухнастіша шерсть і розміри тіла збільшуються згідно з правилом Бергмана [правило Бергмана – закономірність зміни розмірів тварини з огляду на зміни температури]; коли тварина починає полювати на більшу здобич, у неї збільшуються щелепи і кінцівки (щоби швидко бігати й успішніше нападати). Усі ці аспекти адаптації очевидні, але, на мою думку, інтеграція і модульність – це теж адаптації. Якщо для виживання необхідне узгоджене функціонування різноманітних структур, то зростає кореляція між ними, зростає рівень інтеграції. Якщо структура втрачає своє значення, то, навпаки, рівень її кореляції з іншими структурами зменшується, бо зникає потреба в цьому. Відповідно, кожна структура має свій оптимальний рівень інтеграції. Якщо він зменшиться – структура функціонуватиме гірше. Водночас, зростання рівня інтеграції погіршує перспективи подальшої еволюції, бо виду складніше буде еволюціонувати в усіх напрямах, крім напряму лінії найменшого еволюційного опору. Тобто висока інтегрованість може дорого коштувати в еволюційній перспективі (хоча природний добір не дивиться у майбутнє, він діє тільки залежно від сучасної ситуації). Тому зазвичай рівень інтеграції тримається на якомусь оптимальному рівні, необхідному для виду. Що ми й бачимо: у великої нічниці, котра, полюючи на турунів (а це доволі великі комахи з доволі твердими покривами), користується передусім щелепами, – вищий рівень інтеграції черепа та щелепного апарату, ніж у ставкової нічниці, яка ловить водних комах (чи навіть мальків риб) за допомогою своїх задніх кінцівок.
…Є дуже стара і вже класична робота американських дослідників Роберта Бейдера і Джона Голла, присвячена аналізові мінливості і системи кореляції крила кажанів зокрема і їхніх кінцівок узагалі на прикладі двох американських видів нічниць. Основу крила кажанів становить літальна перетинка, натягнута на кістки п’ястку та фаланги пальців із другого по п’ятий; тобто крило кажанів – це, згрубша, долоня. Перший палець не входить до складу крила і лишається окремим.

Крило кажана. Фото Жиля Сан Мартена. Джерело: www.flickr.com/photos/sanmartin/
– Він схожий на гачок.
– Так. І слугує для локомоції по субстрату – щоб прикріпитися до дерева чи до склепіння печери. Чи по землі ходити, як новозеландські містацини. …Так от, Бейдер і Голл показали, що пальці кажана, які пов’язані з крилом, є дуже високоінтегрованими між собою. Але з першим пальцем у них значно нижча кореляція, бо він виконує іншу функцію – теж важливу, але таку, що не потребує значної інтеграції з іншими пальцями. Основну роль у функціонуванні крила відіграють третій, четвертий і п’ятий пальці, їхня мінливість низька, а кореляції між їхніми елементами високі. А в другого пальця мінливість значно більша, ніж у інших пальців, бо функціональне навантаження на цей палець менше, ніж на інші пальці.
Схожі закономірності ми бачимо в багатьох інших випадках. У структур із високим функціональним навантаженням часто низька мінливість і високі кореляції між елементами. Послаблення функції зазвичай призводить до зменшення кореляцій та зростання мінливості. Зокрема, від рудиментарних органів, які втрачають чи майже втрачають свою функцію, можна очікувати, крім зменшення кореляції з іншими гомологічними [побудованими за єдиним планом] і функціонально пов’язаними структурами, ще й дуже великого зростання мінливості.
– А у кажанів теж є рудименти?
– Так.
– Які, наприклад?
– Зараз усе розповім.
– Пане Ігоре, поки ми не заглибились у цю тему, дозвольте запитати про математичний апарат, який ви з колегами використовували. Чи є перспективи для інтеграції технологій штучного інтелекту у ці дослідження?
– Так, гадаю, це цілком можливо. Річ у тім, що штучний інтелект – це, зокрема, і машинне навчання, яке, по суті, використовує чимало методів сучасної статистики: закладаємо певну інформацію і на її основі вдосконалюємо той чи той алгоритм. Зрозуміло, що далі ці технології розвиватимуться і вдосконалюватимуться. Але загалом не бачу нічого екзотичного у застосуванні математики для прогнозування. Навпаки, а як взагалі можна прогнозувати без статистики?
А тепер – про наступні наші наукові результати. Якщо до того йшлося про вивчення черепа, то найсвіжіша наша стаття містить підсумки дослідження зубного апарату нічниць, який має дуже велике значення для них, як і для більшості інших ссавців.
Північноамериканська нічниця Myotis septentrionalis. Фото Джесіки Олендорф. Джерело: animaldiversity.org
Я завжди кажу, що теріологам [дослідникам ссавців] у певному сенсі дуже пощастило: структура, яка є чи не найінформативнішою (і тому їх цікавить найбільше), є найтвердішою в організмі. Дуже багато інформації про ссавців вдалося дізнатися саме завдяки аналізу зубів. Крім того, зуби гомологічні між собою, і можна визначити, які з них мають велике, які – менше функціональне навантаження, а які взагалі рудиментарні. Наприклад, у сучасної людини рудиментарним є третій моляр, або ж так званий зуб мудрості. В індивідуальному розвитку він закладається значно пізніше, ніж інші зуби, часто хворіє, має збільшену мінливість і взагалі створює нам купу проблем. Те саме – з апендиксом. Взагалі будь-який рудимент має кілька характерних рис: спрощену будову, збільшену мінливість, зменшену чи взагалі втрачену функцію, – і наявний у всіх особин виду (бо якщо у більшості особин він ще й повністю втрачається, то це вже не просто рудимент, а атавізм). До речі, зуби мудрості стали рудиментом тільки у сапієнсів – через скорочення зубного апарату. А у неандертальців це були ще цілком функціональні зуби.
Усе сказане стосується і нічниць. У них теж є рудименти. Розгляньмо спершу ставкову нічницю, ту саму, що літає над великими водоймами і полює на личинок комарів. Як на кажана, то вона порівняно велика, але типові комахоїдні ссавці – дрібні, максимум завбільшки з їжака, а літуни – ще менші. Виняток – мурахоїди та панголіни, бо вони спеціалізуються на суспільних комахах, а біомаса термітників чи мурашників цілком здатна прогодувати великого звіра.

Нижня щелепа і череп ставкової нічниці з колекції Ігоря Дзеверіна. Фото пресслужби НАН України
То яка ж зубна система у ставкової нічниці? В принципі, її схема зубів – та сама, що й у нас, людей. Є різці, ікла, премоляри [малі корінні зуби, розташовані між іклами та молярами] і моляри [великі корінні зуби]. У кажанів найкраще функціонально розвинуті зуби – це моляри, бо вони потрібні для подрібнення покривів комах. Тож і мінливість у них – найменша. Проте нас цікавили премоляри. Премолярів у нічниць – три. За порівняльною анатомічною номенклатурою, це другі, треті й четверті премоляри. Перші ж моляри втрачені в усіх кажанів. До речі, їх немає навіть в оніхоніктеріс. Першим плацентним ссавцям була властива зубна формула «3–1–4–3», тобто три різці, одне ікло, чотири премоляри і три моляри. Далі кількість зубів почала скорочуватися. Хоча у деяких видів вона зберіглася незмінною (як, наприклад, у свиней). Зубна формула сучасної людини – «2–1–2–3». На верхній щелепі нічниць – «2–1–3–3», на нижній – «3–1–3–3». Але верхні треті премоляри вже рудиментарні: вони зменшуються (хоч наразі вони ще доволі великі, порівняно з іншими), часто зсуваються в бік від лінії зубного ряду і мають збільшену мінливість. Це класичний приклад рудиментарного зуба. До речі, у більшості кажанів, зокрема в усіх українських кажанів, крім нічниць, верхній третій премоляр узагалі втрачається – лишаються тільки два верхні моляри чи менше. Далі починається рудиментація інших зубів, і наступним втрачається третій нижній моляр. У нічниць він іще абсолютно нормальний, а от у деяких інших кажанів уже теж стає рудиментарним. Потім починають втрачатися деякі моляри і навіть різці. Причому ця втрата відбувається завжди практично за одним і тим самим порядком. Тобто на основі того, які зуби втрачено, можна побудувати послідовність родів кажанів. Ця інформація є у нашій наступній статті. Але це не головний наш результат. Послідовність побудовано, фактично, завдяки літературним даним. Вона передусім демонструє єдиний для гладконосих кажанів механізм морфогенезу [виникнення й розвитку] зубів.
Тепер про наступного нашого персонажа – гостровуху нічницю. Оскільки вона більша за розміром, то навіть візуально легко помітити, які малі у неї рудиментарні зуби, на відміну від добре спеціалізованих молярів, особливо перших двох. Дослідження одразу кількох видів одного роду дало нам змогу надійно порівняти ознаки за функціональною значущістю. Як я вже сказав, це зручно робити, використовуючи серію гомологічних структур, частина яких повністю функціональні, а решта – рудиментарні. Зуби ідеально підходять для такого аналізу. Хоча загалом годяться будь-які серіально гомологічні структури на кшталт хребців хребта.

Нижня щелепа і череп гостровухої нічниці з колекції Ігоря Дзеверіна. Фото пресслужби НАН України
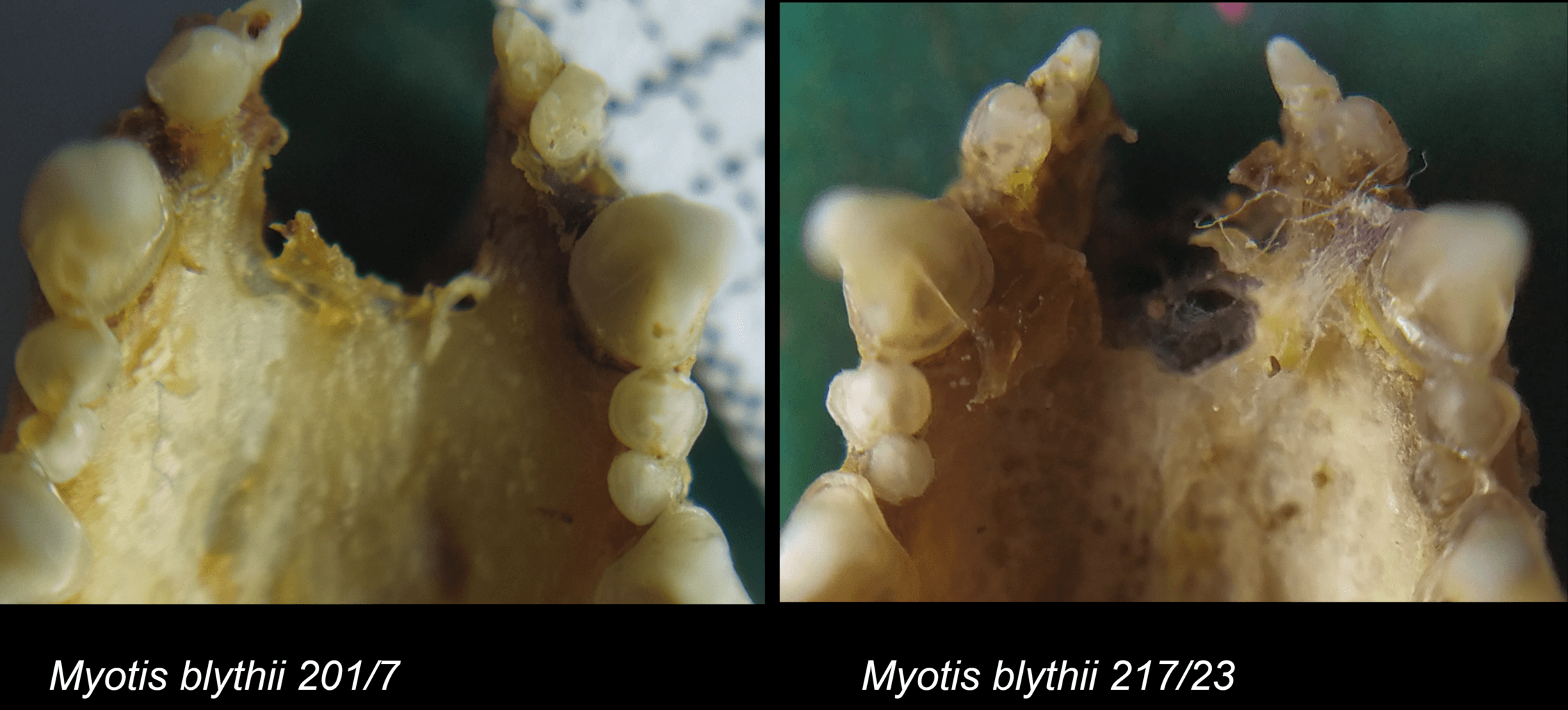
Знімок рудиментарних зубів у нічниць. Авторка світлини – Марія Ґхазалі
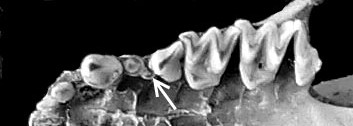
Зуби нічниці (на рудиментарний вказує стрілочка). Фото – з докторської дисертації Ігоря Дзеверіна «Механізми трансформації структур черепа в еволюції нічниць та споріднених груп гладконосих рукокрилих»
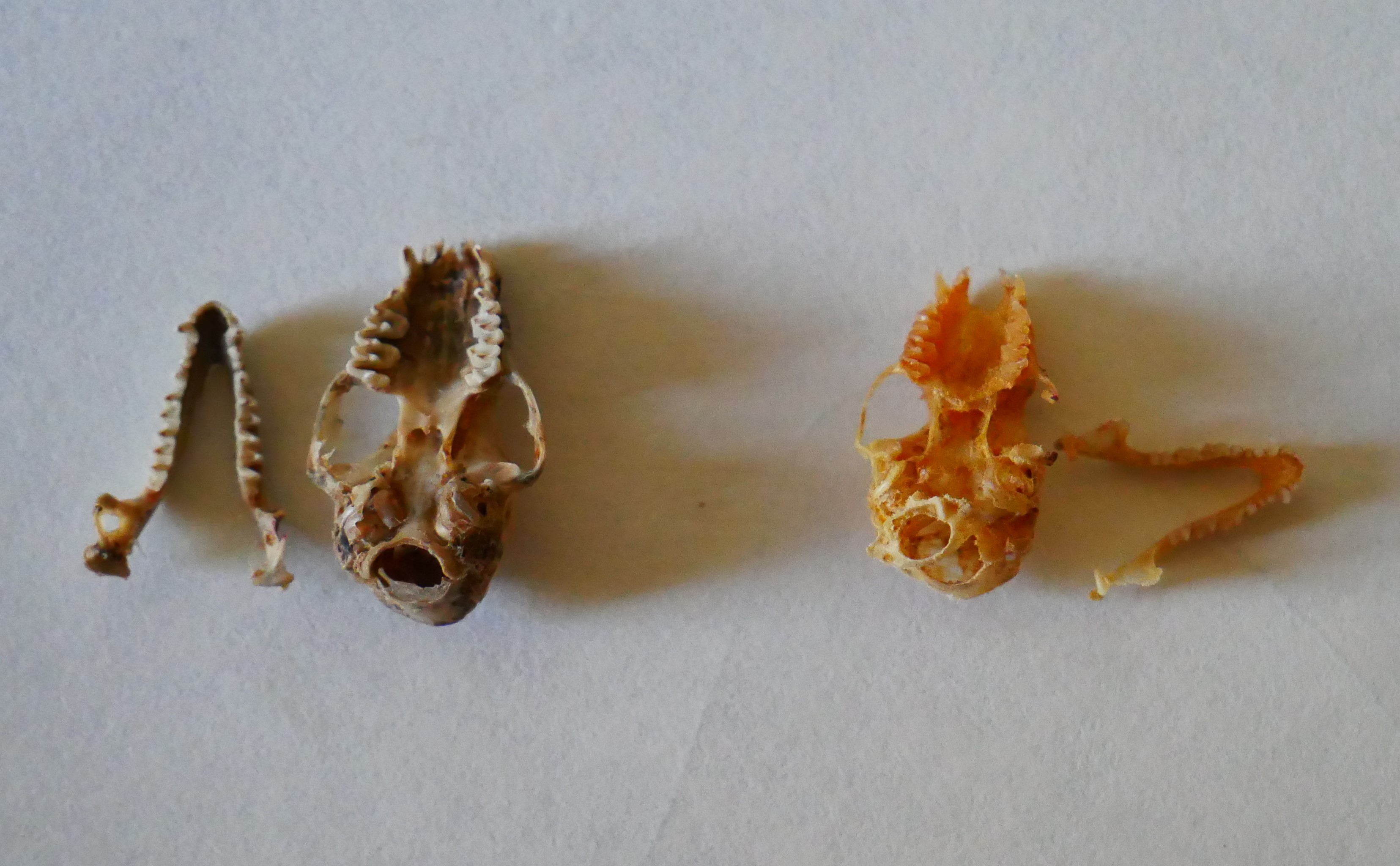
Порівняння щелеп і черепів гостровухої (ліворуч) і ставкової (праворуч) нічниць. Фото пресслужби НАН України
Крім того, тут теж хотілося визначити інтеграцію, подивитись, як зубні структури відповідають на добір, тобто окремо дослідити на зубах те, що вже було досліджено на черепі загалом. Бо, слід сказати, зуби хоч і є частиною черепа, а проте вони трохи самі по собі. Вони мають інший морфогенез, розвиваються за дещо іншими закономірностями. Це очевидно для кожного науковця, який працював із черепом ссавців, і всі ці особливості й відмінності докладно описано у багатьох спеціальних статтях.
Тобто наступною нашою ідеєю було дослідити інтеграцію та модульність саме зубної системи. Найкращий і фактично єдиний фахівець з еволюції зубної системи нічниць в Україні – Марія Гхазалі. Вона має велику базу даних, великий досвід і присвятила цій темі свою кандидатську дисертацію [тема дисертації Марії Гхазалі на здобуття ступеня кандидата біологічних наук – «Одонтометрична мінливість нічниць, Myotis (Chiroptera, Vespertilionidae), фауни України: еволюційні, функціональні, екологічні аспекти» (2010)]. Тому я запропонував об’єднати наші зусилля – і далі ми працювали вже втрьох: Марія Гхазалі, Оксана Верцімаха і я. Довелося дещо модифікувати методику пошуку структури зв’язків. Вагомий внесок у це зробила Оксана. Внесок Марії теж дуже вагомий, починаючи з того, що ми працювали із даними, які зібрала саме вона. Крім того, Марія запропонувала багато цікавих ідей. У цій роботі ми зосередилися тільки на двох таксономічних одиницях – великій нічниці та європейській гостровухій нічниці, бо про інших, на жаль, поки що немає достатньої кількості даних.
Що ж у нас вийшло? По-перше, виявилося, що найбільш інтегровані і найтісніші кореляційні зв’язки мають ті ознаки, які забезпечують оклюзію зубів, тобто правильний прикус. Це передбачувано, та все ж варто було перевірити. Інші ознаки варіюють більш вільно. Далі. Ми дуже чітко зафіксували надзвичайно високу мінливість рудиментарного зуба – вже згаданого великого третього премоляра. Для візуалізації мінливості у морфометрії заведено застосовувати такий показник, як коефіцієнт варіації. Це відношення стандартного відхилення до середнього значення ознаки. З погляду високої математики, коефіцієнт варіації – можливо, недосконалий інструмент, але він дуже корисний у морфометричних дослідженнях і широко застосовується в них. Для нашого дослідження він також виявився дуже корисним і добре показав, як зростає мінливість рудиментарної структури. Одночасно зі зростанням мінливості зменшується кореляція рудиментарного зуба з іншими зубами. Все як і очікувалося. Ми також побачили, що у великої нічниці інтеграція щонайшвидше вища, ніж у гостровухої нічниці, та продемонстрували дуже високий рівень онтогенетичних обмежень еволюційного процесу в цих видів. Тобто у підсумку вивчення зубної системи нічниць ми одержали картину, якісно схожу з тим, що ми побачили на черепі, але у деталях вона все ж відрізняється. Ну, і знов-таки, знову підтвердилася думка про те, що і інтеграцію, і модульність можна розглядати як адаптації самі по собі. Якщо є еволюційна необхідність забезпечити злагоджене функціонування великої кількості структур, то їхня інтеграція зростає, вони об’єднуються в єдиний модуль, інтеграція всередині якого теж зростає. Якщо ж якась структура втрачає свою цінність, то її кореляції з іншими ознаками зменшуються, вона випадає з модуля, її мінливість зростає, і, якщо це триватиме далі, то ознака перетворюється на рудимент, потім – на атавізм і зрештою зникає взагалі. Як верхні треті премоляри у більшості кажанів.
– Тобто рудимент приречений зникнути?
– Ні. Він зникне з високою імовірністю, але буває по-всякому. Іноді, навпаки, в процесі подальшої еволюції рудимент знову починає збільшуватись, якщо знайде собі функцію й увійде до якогось нового модуля.
Повернімося до рибоїдної нічниці – американського аналога нашої ставкової нічниці, щоправда більшого за розміром і спеціалізованого на полюванні риби. Ми згадували її на початку розмови. У рибоїдної нічниці верхні треті премоляри почали вторинно збільшуватися. І коли 100 років тому дослідники це помітили, то навіть описали особин зі збільшеними рудиментами як окремий від нічниці рід, назвавши його пізоніксом. Пізніше за допомогою генетичних методів було доведено, що це звичайна нічниця, тільки з дещо іншими еволюційними трендами, тому її повернули до роду нічниць, і її правильна назва – рибоїдна нічниця (Myotis vivesi). А назву «пізонікс», за правилами зоологічної номенклатури, тепер застосовують до американського підроду нічниць. Отже, на прикладі верхніх третіх молярів рибоїдної нічниці, ми бачимо, як рудимент знову набуває цінності і починає прогресивно еволюціонувати.

Рибоїдна нічниця. Добре видно відносно збільшені верхні треті премоляри. Фото Скотта Трейджсера. Джерело: https://www.flickr.com/photos/naturestills/
Наведу ще один приклад, і такий, що стосується значно масштабнішого еволюційного перетворення. Еволюція наземних хребетних – від земноводних до плазунів, птахів і ссавців – була шляхом до унезалежнення від водного середовища. Це, зокрема, означало втрату шкірних залоз і набуття сухої шкіри. Так сталось і з плазунами, і з птахами. Ссавці є нащадками дуже примітивних плазунів, настільки близьких до земноводних, що деякі дослідники припускали навіть, ніби ссавці походять безпосередньо від земноводних, оминаючи плазунову еволюційну стадію. Це перебільшення, але якась частина істини у ньому є. Так от: предок ссавців був настільки примітивним плазуном, що у нього зберігалися шкірні залози. Рудиментарні, звісно. Але у ссавців ці рудименти почали прогресивно розвиватись і дали початок потовим, сальним і молочним залозам. Тобто те, що було рудиментом, втраченим у плазунів і птахів, дало у ссавців початок прогресивному розвиткові. І не просто прогресивному розвиткові: ці залози – важлива частина ароморфозу ссавців, тоді як ароморфоз плазунів був пов’язаний, навпаки, із втратою цих залоз. Отже, нова еволюція рудиментів трапляється в еволюції, хоч і не часто.
Схема еволюції ссавців. Автор – Пітер Бішоп. Джерело: phys.org
Для еволюційної новації взагалі не принципово, звідки береться її джерело. Це може бути абсолютно новий морфогенез на базі якихось мутацій, напіввтрачений рудимент або ще щось. Старі автори виокремлювали так звані орименти – зачатки нових органів. Можна сказати, що маленькі ріжки у жирафи – це оримент високорозвинених рогів, таких як у оленів чи у порожнисторогих [порожнисторогі – жуйні парнокопитні ссавці, роги яких є нерозгалуженими кістковими виростами, вкритими кератиновим футляром; до цієї родини належать, зокрема, антилопи, бізони, бики, вівці, кози].

Жираф. Фото: Getty Images

Американський білохвостий олень. Фото: www.britannica.com
Із усіх цих прикладів випливає, що для еволюції великої різниці між рудиментами і ориментами немає, а рудимент іноді може стати ориментом. Так само у процесі еволюції ознака може перейти з одного модуля до іншого, якісь ознаки можуть об’єднатись у новий модуль і дати початок чомусь принципово новому. Взагалі-то в еволюції дуже рідко з’являється щось принципово нове, але – з’являється. Приклад такого зовсім нового – мозолисте тіло між півкулями головного мозку у плацентних ссавців, яке забезпечує зв’язок між процесами, що відбуваються в цих півкулях, і, очевидно, сприяє нервовій діяльності. Цієї еволюційної новації немає навіть у сумчастих. Щоб щось принципово нове з’явилося, потрібно, щоб гени, умовно кажучи, об’єдналися в нову систему регуляції та почали разом працювати, що, своєю чергою, призведе до появи і нових фенотипних ознак. Це ідея і тема багатьох досліджень Ваґнера зі співавторами [Ґюнтер Ваґнер – австрійський та американський еволюційний біолог, професор Єльського університету]. Колись, 12 років тому, я опублікував огляд цих концепцій.
Отже, ми у своєму дослідженні зафіксували динаміку ознак між модулями. Залежно від необхідності пристосуватися до певних умов середовища, інтеграція ознак зростає або зменшується. І там, де це потрібно, кореляційні системи перебудовуються, виникають нові модулі й зникають старі.
До речі, ми стикнулися з проблемою, що наукова термінологія у цій галузі ще не усталена. Зокрема, ми вживаємо терміни «адаптивні перебудови», «перебудови кореляційних систем» і аналогічні, бо ними послуговувався Іван Шмальгаузен [засновник Інституту зоології і його директор у 1930–1941 роках, академік], коли описував еволюцію кореляції (у нього йшлося про «адаптивні перебудови організму»). Англійською теж можливі різні варіанти – restructuring (дослівно – реструктуризація), repatterning (зміна шаблонів). Для своєї статті ми з колегами обрали термін “restructuring”, взоруючись на французьке “restructuration”.
Ось такі результати ми здобули.
– Що плануєте досліджувати далі, щоб остаточно обґрунтувати і довести свою гіпотезу?
– Хочу збільшити масштаб і проаналізувати еволюцію кореляційної системи на ширшій вибірці видів. Бо ми вивчали, нагадаю, шість таксономічних одиниць, п’ять із яких є дуже близькими родичами, а цікаво дізнатись, як ці процеси відбуваються в масштабах усього дуже різноманітного роду. Або й в інших родах. Сподіваюся, що зможемо зібрати дані і виконати цю роботу. Принаймні спробуємо. Бо це доволі перспективний напрям і широке поле – дослідження того, як кореляційні системи перебудовуються в різних еволюційних лініях і що це дає з погляду еволюції.

Водяна нічниця. Фото Жиля Сан Мартена. Джерело: www.flickr.com/photos/sanmartin/
– Кажани, як і китоподібні, вразливі тварини, інакше вони не мали б охоронного статусу. Чи можуть ваші з колегами дослідження допомогти захистити їх? Іншими словами, чи матимуть ваші результати, крім впливу на розвиток науки, ще й якийсь практичний ефект?
– Складно сказати. Прямого прикладного ефекту, мабуть, не буде. Це все ж таки дослідження еволюції. Хоча вони допомагають краще зрозуміти, скажімо, як кажани сформувалися, де їхні вразливі місця, як вони відповідатимуть на зміни напряму добору і так далі. Тобто певне застосування можливе, але – непряме.
Я зараз знов відхилюся від теми, щоб пояснити свою думку. Теоретичний фундамент наших досліджень – це методи еволюційної кількісної генетики, до розроблення яких суттєво доклався американський дослідник Рассел Ланде [Рассел Ланде – американський еволюційний біолог і еколог, нині професор Норвезького науково-технологічного університету]. У своїх теоретичних роботах він спирався переважно на концепції полігенно детермінованих ознак і показав, як ці ознаки еволюціонують. Але були у Ланде пізніше і прикладні дослідження – екологічні, демографічні. Зокрема, одну з праць він присвятив плямистій сові (Strix occidentalis caurina). Це вид, що мешкав на західному узбережжі США – у штатах Орегон і Вашингтон. Чому плямиста сова зацікавила Ланде? Вона почала вимирати. Виявилося, що це вибагливий вид – для нормального життя йому потрібні саме старі дуплясті дерева. Тобто можна висадити новий ліс замість вирубаного старого, але радіти буде зарано, бо в цих новонасаджених деревах сова не житиме – вона заселяє тільки дерева на заключному етапі сукцесії. І от Ланде застосував математичний апарат, щоб з’ясувати, які чинники найбільше впливають на демографію плямистої сови. Це надзвичайно талановита робота, як, утім, і решта його робіт. І ця робота саме прикладна.

Плямиста сова. Фото Джареда Г’юї. Джерело: www.flickr.com/photos/jaredhughey
Те, що робимо ми, не є прикладним у сенсі цього дослідження Ланде. Наша робота ближча до його теоретичних праць: ми залучаємо до емпіричного матеріалу розроблені ним та іншими авторами теоретичні моделі та методики. Та все ж я сподіваюся, що краще розуміння того, на чому ґрунтується цілісність організації організму кажанів (а саме – оцінки рівнів мінливості й рівнів інтеграції), допоможе розробити якісь прикладні програми, зокрема і для їх охорони. Бо, коли порушуються умови середовища, ця цілісність теж порушується. Але я не хотів би, щоб мене зрозуміли неправильно. Повторюю, зв’язок між нашими дослідженнями і практичним ефектом – непрямий. Ніколи не треба вигадувати практичне значення там, де є тільки теоретичне. Мета науки – з’ясувати закони природи і запропонувати загальну картину світу. Хоча, з іншого боку, практичні застосування можливі виключно на основі законів природи, які з’ясувала фундаментальна наука. Річ у тім, що загальна теорія еволюційних процесів може спрацювати в дуже несподіваних аспектах. Наприклад, динаміка будь-якої інвазії – байдуже, бур’янів чи інфекції (навіть того ж ковіду) – описується тими само рівняннями, які розроблено для популяційної генетики. Тож, можливо, наші дослідження інтеграції організму стануть у пригоді, наприклад, для розроблення геронтологічних програм.
– Тобто вже виконані дослідження – необхідна підвалина, крок на цьому шляху?
– Так. Просто про це ще зарано говорити. Ми щойно почали. У цій темі дуже багато проблем. Кажани аномально довго живуть порівняно з іншими ссавцями таких розмірів і мають аномально високу резистентність до різних вірусних і бактеріальних хвороб. Трапляються, звісно, епідемії, смертельно небезпечні для кажанів, наприклад, синдром білого носа. Але для багатьох інших збудників кажани можуть бути безсимптомними носіями. Звідси й розмови про те, що ковідом люди заразилися від кажанів. Так от, прикладна цінність науки може полягати й у тому, щоб довести, що цього не було і бути не могло, що причина пов’язана не з еволюцією, а радше з екологією.
Людина – теж еволюційна аномалія. Так, великі ссавці живуть довше, ніж дрібні, але ми живемо значно довше, ніж могли б жити, зважаючи на наші розміри тіла. Завбільшки ми як шимпанзе і менші, ніж горили, але тривалість нашого життя десь удвічі більша.
– Чи є припущення чому?
– Найочевидніше припущення полягає в тому, що довге життя потрібне було нам для розвитку соціальності. Бо у нас аномально висока тривалість дитинства і дуже суттєву роль відіграє обмін інформацією, носіями якої є старі особини, що мають навчити молоде покоління. Тому інших варіантів не було – ми могли еволюціонувати тільки в бік зростання тривалості життя. Очевидно, цей процес був дуже інтенсивний і досі не завершився. У тому розумінні, що ми як вид стикнулися з побічними результатами, яких добір іще не здолав. Це загалом відомий факт, що доволі інтенсивний добір може призводити до побічних ефектів. Вони особливо помітні у селекційних програмах: добір за однією лиш ознакою знижує життєздатність багатьох порід. Гадаю, що у людей висока ймовірність пошкоджень, пов’язаних із соматичним мутагенезом, зокрема онкозахворювань, є побічним наслідком того, що тривалість життя швидко зросла. Добір проти раку дуже малоінтенсивний – просто через те, що люди встигають розмножитися, перш ніж захворіють на онкопатології. А що буде після розмноження, добір уже не цікавить.
У кажанів дещо інша ситуація: ми можемо припускати, що збільшення тривалості життя – це пряма адаптивна відповідь на низьку плодючість. Дитинча дуже велике порівняно з матір’ю (здається, чи не найбільше новонароджене, якщо порівнювати з розміром матері, серед ссавців узагалі). Самиця народжує зазвичай одне, дуже рідко – двох дитинчат. Середня тривалість покоління, тобто середня відстань між народженням самиці кажана і народженням у неї малят, – приблизно 4 роки. Деякі самиці народжують навіть у віці 10 років. Такі випадки, щоправда, нечисленні, але вони є. І це дуже багато для таких тварин. А отже, щоб нормально відтворюватися, кажани мають розмножуватись якнайдовше. І інших варіантів тут немає. Кажани літають, тому конструкція їхнього тіла, пристосована до забезпечення польоту, дуже жорстка і майже не змінюється. Добір не підтримав би мутації в бік збільшення плодючості.

Малюк водяної нічниці. Фото Жиля Сан Мартена. Джерело: www.flickr.com/photos/sanmartin/
Звісно, у людей також низька плодючість, і ми можемо припускати з високою ймовірністю, що розвиток соціальності в нашій еволюції був спрямований саме до збільшення шансів на виживання дітей, та й дорослих. Однак у людей зростання тривалості життя, радше, було адаптивною відповіддю на зростання значення соціальності, ніж безпосередньо на низьку плодючість.
Цікаво, що у деяких кажанів розвивається соціальність, чимось подібна до нашої. У вампірів, або ж десмодів (тобто невеликої південно- й центральноамериканської групи кажанів, які живляться кров’ю інших тварин), поширене таке явище, як взаємодопомога: кажан, який не зміг знайти жертву, повертається до своєї соціальної групи, де його погодують. Практикують вони і взаємний грумінг. Отакі форми альтруїзму. Хоча з мавпами, звісно, не порівняти. Зрештою, так чи так, а виникають підстави для зростання тривалості життя. Очевидно, кажани і кити в цьому успішніші, ніж люди – можливо, через те, що еволюціонують цим шляхом значно довше. Але ймовірний шлях саме такий, як я описав.
– Словом, кажани – об’єкт цікавий і однозначно вартий уваги.
– Безумовно. Навряд чи я можу сказати інакше, якщо все своє життя їх досліджую (усміхається). Є відомий анекдот про якогось видатного фахівця, який почав вивчати червів іще студентом і займався ними аж до пенсії. Коли його спитали, чи не набридло, він відповів: «Черв’як такий довгий, а життя таке коротке». Зважаючи на те, скільки років я вивчаю нічниць, то мені вже і про себе час розповідати такі анекдоти.

Велика нічниця. Фото Жиля Сан Мартена. Джерело: www.flickr.com/photos/sanmartin/
Спілкувалася Сніжана Мазуренко